Нефрон, состоящий из сосудистого клубочка, его капсулы и почечных канальцев (рис. 1.1), имеет высокую структурно-функциональную специализацию.
Сосудистый клубочек. В каждой почке содержится около 1—1,3 млн клубочков. Сосудистый клубочек имеет около 50 капиллярных петель, между которыми найдены анастомозы, что позволяет клубочку функционировать как «диализирующая система». Стенка капилляра представляет собой клубочковый фильтр, состоящий из эпителия, эндотелия и располагающейся между ними базальной мембраны (БМ).
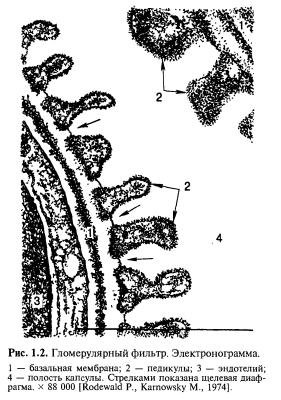
Эпителий клубочка, или подоцит, состоит из крупного клеточного тела с ядром в его основе, митохондриями, пластинчатым комплексом, эндоплазматической сетью, фибриллярными структурами и другими включениями. Большие отростки подоцита отходят из перинуклеарной зоны; они напоминают «подушки», охватывающие значительную поверхность капилляра. Малые отростки, или педикулы, отходят от больших почти перпендикулярно и закрывают собой все свободное от больших отростков пространство капилляра (рис. 1.2 и 1.3). Педикулы тесно прилежат друг к другу, межпедикулярное пространство составляет 25— 30 нм.

Подоциты связаны между собой пучковыми структурами, образующимися из плазмолеммы. Фибриллярные структуры особенно отчетливо выражены между малыми отростками подоцитов, где они образуют так называемую щелевую диафрагму (см. рис. 1.3), которой отводится большая роль в гломерулярной фильтрации. Щелевая диафрагма, имея филаментарное строение (толщина 6 нм, длина 11 нм), образует своеобразную решетку, или систему пор фильтрации, диаметр которых у человека 5—12 нм. Снаружи щелевая диафрагма покрыта гликокаликсом, т.е. сиалопротеиновым слоем цитолеммы подоцита, внутри она граничит с lamina rаrа externa БМ капилляра (рис. 1.4).

На поверхности подоцитов найден эпителиальный антиген почечных канальцев, гликопротеин щеточной каймы (gp 330) и подоксалин. Подоцитарный антиген представлен эндогенным гликопротеином, который включен в состав рецепторов, осуществляющих эндоцитоз в подоцитах; gp 330 локализован в углублениях плазматической мембраны ножек подоцитов и в структурах синтеза этих клеток — эндоплазматической сети и пластинчатом комплексе Гольджи. Подоксалин — основной сиалопротеин клубочка, обеспечивающий отрицательный заряд БМ; локализован в основании ножек подоцитов и на поверхности самих клеток, обращенной к базальной мембране.
Функцию фильтрации осуществляет не только щелевая диафрагма, но и миофиламенты цитоплазмы подоцитов, с помощью которых происходит их сокращение. Так, «субмикроскопические насосы» перекачивают ультрафильтрат плазмы в полость капсулы клубочка. Той же функции транспорта первичной мочи служит и система микротрубочек подоцитов. С подоцитами связана не только функция фильтрации, но и, как уже говорилось, продукция вещества БМ. Кроме того, они экспрессируют антигены, общие с клетками крови. В цистернах гранулярной эндоплазматической сети этих клеток находят материал, аналогичный веществу базальной мембраны.
Изменения подоцитов чаще всего бывают вторичными и обычно наблюдаются при про-теинурии, нефротическом синдроме (НС). Они выражаются в гиперплазии фибриллярных структур клетки, исчезновении педикул, вакуолизации цитоплазмы и нарушении щелевой диафрагмы. Эти изменения связаны как с первичным повреждением базальной мембраны, так и с самой протеинурией. Инициальные и типичные изменения подоцитов в виде исчезновения их отростков характерны для липоидного нефроза.
Эндотелиальные клетки капилляров клубочка имеют поры размером до 100—150 нм и снабжены специальной диафрагмой. Поры занимают около 30 % эндотелиальной выстилки, покрытой гликокаликсом. Поры рассматривают как основной путь ультрафильтрации, но допускают и трансэндотелиальный путь, минующий поры; в пользу этого допущения говорит высокая пиноцитозная активность гломерулярного эндотелия. Помимо ультрафильтрации, эндотелий гломерулярных капилляров участвует в образовании вещества БМ.
Изменения эндотелия капилляров клубочка разнообразны: набухание, вакуолизация, некробиоз, пролиферация и десквамация, однако преобладают деструктивно-пролиферативные изменения, столь характерные для гломерулонефрита (ГН).
Важную роль в регуляции кровотока играет поверхность эндотелиальных клеток. При повреждении этих клеток на их поверхности появляются многочисленные выросты (аркадная формация), создающие турбулентные завихрения кровотока и способствующие адгезии лейкоцитов к эндотелию. Кроме того, эндотелиальные клетки участвуют в регуляции кровотока, секретируя эндотелины, оказывающие сосудосуживающий эффект, и оксид азота (NO), вызывающий вазодилатацию.
Базальная мембрана клубочковых капилляров, в образовании которой участвуют не только подоциты и эндотелий, но и мезангиальные клетки, имеет толщину 250—400 нм и в электронном микроскопе выглядит трехслойной: центральный плотный слой (lamina densa) окружен более тонкими слоями с наружной (lamina rаrа externa) и внутренней (lamina гага interna) стороны. Собственно БМ служит lamina densa; наружный и внутренний слои являются по существу гликокаликсом подоцитов и эндотелия. Филаменты lamina densa толщиной 1,2—2,5 нм входят в «подвижные» соединения с молекулами окружающих их веществ и образуют тиксотропный гель.
В состав БМ входит несколько специфических белков, которые объединяются в высокоорганизованный матрикс. Это прежде всего коллаген IV типа, ламинин и 2 формы гепарансульфатпротеогликанов (ГСПГ), которые образуют «рабочую сеть», соединяющуюся с клетками и другими компонентами матрикса, включая коллаген V типа, энтактин, ВМ-40, нефритогенозид, фибронектин. Суммируя многочисленные данные о распределении структурных компонентов ГБМ, можно сказать, что для коллагена IV типа характерна локализация во всех слоях БМ, но наиболее высокая концентрация его найдена во внутренней части lamina densa. Ламинин обнаружен во всех слоях БМ, наиболее высокая концентрация характерна для lamina densa externa и lamina rаrа intema; там же выявлены в высокой концентрации и ГСПГ. Энтактин, ВМ-40, нефритогенозид найдены в lamina rаrа interna, фибронектин и коллаген V типа — как в lamina densa, так и в окружающих ее пластинках.
Если с коллагеновыми белками связано структурирование БМ, то с неколлагеновыми гликопротеинами — ламинином, энтактином и фибронектином — адгезия клеток с базальной мембраной. С возрастом количество коллагена БМ значительно увеличивается, а гликопротеинов — снижается. Протеингликаны БМ совместно с сиалопротеинами подоцитов и в меньшей мере эндотелия обеспечивают отрицательный заряд базальной мембраны, который является основным барьером для анионных и нейтральных макромолекул.
С присутствием в плотной пластинке коллагеноподобных филаментов связана гипотеза о порах фильтрации в базальной мембране. Показано, что средний радиус пор мембраны равен 2,9 нм и определяется расстоянием между нормально расположенными и неизмененными филаментами коллагеноподобного белка. При падении гидростатического давления в капиллярах клубочков первоначальная «упаковка» коллагеноподобных филаментов в БМ изменяется, что ведет к увеличению размера пор фильтрации. Предполагают, что при нормальном кровотоке поры базальной мембраны гломерулярного фильтра достаточно велики и могут пропускать молекулы альбумина, IgG, каталазы, но проникновение этих веществ ограничено высокой скоростью фильтрации. Фильтрация ограничена также дополнительным барьером гликопротеинов (гликокаликс) между мембраной и эндотелием, причем этот барьер в условиях нарушенной гломерулярной гемодинамики повреждается.
Для поддержания нормальной клубочковой фильтрации большое значение имеет отрицательный заряд стенки капилляров клубочков. Благодаря отрицательному заряду БМ и глико-каликсовой оболочки, покрывающей подоциты, от стенки капилляра отталкиваются белковые молекулы плазмы, которые при физиологических значениях pH имеют отрицательный заряд. Белки плазмы поэтому не проходят дальше субэндотелиального слоя БМ, но для тех молекул, которые прошли ее, последним барьером является щелевая диафрагма. Инициальными моментами в возникновении протеину-рии служат очаговые дефекты гломерулярной БМ (микроперфорации, очаговое оголение подоцитов). Через такие очаговые дефекты белки выходят в полость капсулы, что в свою очередь изменяет первоначальный заряд стенки капилляра, снимает часть отрицательного заряда. Это приводит к усилению фильтрации белков через гломерулярный фильтр и появлению про-теинурии.
Изменения БМ клубочка характеризуются ее утолщением, гомогенизацией, разрыхлением и фибриллярностью. Утолщение БМ встречается при многих заболеваниях с протеинурией. При этом наблюдаются увеличение промежутков между филаментами мембраны и деполимеризация цементирующего вещества, с чем связывают повышенную порозность мембраны для белков плазмы крови. Кроме того, к утолщению БМ гломерул ведут мембранозная трансформация, в основе которой лежит избыточная продукция вещества БМ подоцитами, и мезангиальная интерпозиция, представленная «выселением» отростков мезангиоцитов на периферию капиллярных петель, отслаивающих эндотелий от БМ.
При многих заболеваниях с протеинурией, помимо утолщения мембраны, методом электронной микроскопии выявляются различные отложения (депозиты) в мембране или в непосредственной близости от нее. При этом каждому отложению той или иной химической природы (иммунные комплексы, амилоид, гиалин) соответствует своя ультраструктура. Наиболее часто в БМ выявляются депозиты иммунных комплексов, что ведет не только к глубоким изменениям самой мембраны, но и к деструкции подоцитов, гиперплазии эндотелиальных и мезангиальных клеток.
Капиллярные петли связывает друг с другом и подвешивает наподобие брыжейки к гломерулярному полюсу соединительная ткань клубочка, или мезангий (рис. 1.5), структура которого подчинена в основном функции фильтрации и репарации при повреждении мембраны. В мезангии различают мезангиоциты (мезангиальные клетки) и мезангиальный матрикс.

В мезангиоцитах хорошо выражены пластинчатый комплекс, гранулярная эндоплазматичес-кая сеть; в них много мелких митохондрий, рибосом, лизосом. Цитоплазма клеток богата основными и кислыми белками, тирозином, триптофаном, гистидином, полисахаридами, РНК, гликогеном. Своеобразие ультраструктуры и богатство пластинчатого материала обусловливают высокие секреторные и гиперпластические потенции мезангиальных клеток. Выделяют 3 типа мезангиоцитов: гладкомышечного типа (они преобладают); костномозгового происхождения (относятся к системе мононуклеарных фагоцитов); транзиторные типа моноцитов. Мезангиоциты гладкомышечного типа осуществляют синтез всех компонентов мезангиального матрикса и их утилизацию, костномозговые мезангиоциты заведуют функцией фагоцитоза различных макромолекул, в том числе циркулирующих иммунных комплексов, контролируют синтетическую функцию гладкомышечных мезангиальных клеток, выступают в роли Ia-положительных (антигенпредставляющих) клеток.
Основными компонентами мезангиального матрикса являются коллагены IV и V типов — ламинин и фибронектин. В отличие от БМ матрикс не содержит энтактина и других «мелких» гликопротеинов, вместо ГСПГ присутствует хондроитинсульфатпротеогликаны (ХСПГ). В функциональном отношении мезангиальный матрикс, образуя систему каналов, выполняет роль «мусоропровода» для анионных и нейтральных макромолекул, микроорганизмов и т. д.
Мезангиоциты принимают активное участие в иммунных реакциях (la-положительные клетки). Они способны реагировать на те или иные повреждения гломерулярного фильтра продукцией вещества БМ, в чем проявляется репаративная реакция в отношении основного компонента гломерулярного фильтра. Гипертрофия и гиперплазия мезангиальных клеток ведут к расширению мезангиума, к его интерпозиции, когда отростки клеток, окруженные мембраноподобным веществом, или сами клетки «выселяются» на периферию клубочка, что вызывает утолщение и склероз стенки капилляра, а в случае прорыва эндотелиальной выстилки — облитерацию его просвета. С интерпозицией мезангия связано развитие гломерулосклероза при многих гломерулопатиях (ГH, диабетический и печеночный гломерулосклероз и т.д.). Мезангиоциты выполняют и фагоцитарную функцию — очищение клубочка, его соединительной ткани.
Мезангиоциты являются основным источником медиаторов, регулирующих как тонус капиллярных петель (оксид азота), так и пролиферацию клеток почечного клубочка (ИЛ-1, ФРФ, ТцФР), и синтез ими компонентов внеклеточного матрикса (ТФРЬ, металлопротеиназы, ингибиторы металлопротеиназ).
Мезангиальные клетки как один из компонентов юкстагломерулярного аппарата (ЮГА) способны в определенных условиях к инкреции ренина. Этой функции служат, видимо, взаимоотношения отростков мезангиоцитов с элементами гломерулярного фильтра: определенное количество отростков перфорирует эндотелий клубочковых капилляров, проникает в их просвет и имеет непосредственные контакты с кровью. Считают, что мезангиоциты способны к сокращению, которое подчинено фильтрационной функции. В цитоплазме мезангиальных клеток найдены фибриллы, обладающие актиновой и миозиновой активностью.
Получены данные о механизмах взаимодействия клеток почечного клубочка. Так, подоциты в норме секретируют гепариноподобное вещество, ингибирующее пролиферацию мезангиоцитов, при деструкции подоцитов образуется МАК — мембраноатакующий комплекс, с чем связывают развитие сегментарного гиалиноза клубочков. Мезангиоциты, подоциты и эндотелиоциты клубочка секретируют метаболиты арахидоновой кислоты (простагландины, лейкотриены, тромбоксаны), которые регулируют тонус этих клеток, почечный кровоток и иммунные реакции в клубочках.
Капсула клубочка представлена БМ и эпителием. Мембрана, продолжающаяся в главный отдел канальцев, состоит из ретикулярных волокон. Тонкие коллагеновые волокна закрепляют клубочек в интерстиции. Эпителиальные клетки фиксированы на БМ с помощью филаментов, содержащих актомиозин. На этом основании эпителий капсулы рассматривают как разновидность миоэпителия, изменяющего объем капсулы, что способствует фильтрации. Эпителий имеет кубическую форму, но в функциональном отношении близок к эпителию главного отдела канальцев; в области полюса клубочка эпителий капсулы переходит в подоциты.







 23.01.2018
23.01.2018