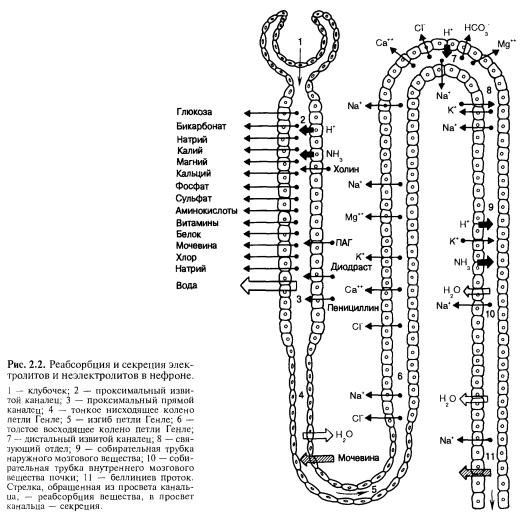
Натрий и хлор. Эти ионы преобладают в молярном отношении среди различных неорганических и органических компонентов плазмы крови и внеклеточной жидкости, их доля наиболее высока среди осмотически активных веществ ультрафильтрата. У взрослого человека концентрация ионов натрия в плазме крови составляет около 143 ммоль*л-1, ионов хлора — 105 ммоль*л-1. В суточной моче концентрация натрия обычно колеблется от 40 до 220 ммоль*л-1 в отдельных пробах мочи она может значительно снижаться. Широкий диапазон колебаний содержания ионов натрия и хлора в моче характерен для нормально функционирующей почки. На примере описания всасывания этих ионов в различных отделах нефрона удобно охарактеризовать общие механизмы функционирования отдельных частей почечных канальцев.
В проксимальном сегменте нефрона всасывается около 2/3 профильтровавшейся воды, натрия, хлора, практически полностью реабсорбируются аминокислоты, глюкоза, витамины, белки и многие другие вещества (рис. 2.2). Этот сегмент нефрона, особенно его прямой отдел, имеет важное значение в секреции из крови в просвет канальца органических кислот и оснований, в самых дистальных частях нефрона и собирательных трубках секретируются ионы калия. Следует отметить, что термин «секреция» имеет два значения применительно к функциям почки: перенос веществ в неизмененном виде через клетки канальца из крови в его просвет и секреция в содержимое канальца или в кровь вновь синтезированных веществ. Наиболее часто термин «секреция» в связи с процессом мочеобразования используют в первом значении этого слова. К секретируемым в почке веществам относятся, например, диодраст, феноловый красный, пенициллин, калий, ионы водорода. В отделах почечных канальцев, следующих за прямым проксимальным сегментом нефрона, всасываются главным образом вода и ионы.

В проксимальном сегменте нефрона всасывается большая часть натрия, но его концентрация в канальцевой жидкости остается такой же, как в плазме крови. Особенность проксимальной реабсорбции заключается в том, что натрий и другие реабсорбируемые вещества всасываются с осмотически эквивалентным объемом воды и содержимое канальца всегда остается изоосмотичным плазме крови. Это обусловлено высокой проницаемостью для воды стенки проксимального канальца.
В начальных отделах канальца главным анионом, сопровождающим натрий, является бикарбонат. Стенка этой части нефрона для хлоридов малопроницаема. Это приводит к постепенному увеличению концентрации ионов хлора, которая возрастает в 1,4 раза по сравнению с плазмой крови. В начальных частях проксимального канальца интенсивно реабсорбируются глюкоза, аминокислоты и некоторые другие органические компоненты ультрафильтрата. Таким образом, в конечных частях проксимального извитого канальца состав изоосмотической жидкости существенно изменяется — из нее всасываются основная масса бикарбоната, многие органические вещества, но концентрация ионов хлора становится выше. Оказалось, что в прямой части проксимального канальца в отличие от извитой части имеется высокая проницаемость межклеточных контактов для хлоридов. В связи с тем что их концентрация в просвете выше, чем в околоканальцевой жидкости и в крови, они начинают по градиенту пассивно реабсорбироваться из канальца, увлекая за собой натрий и воду. В прямом отделе проксимального канальца происходит как активный транспорт натрия, так и пассивная реабсорбция хлоридов и движение части натрия вместе с ними по межклеточным промежуткам, обладающим высокой проницаемостью для хлоридов.
Резюмируя сказанное о реабсорбции натрия и хлоридов в проксимальных канальцах, следует рассмотреть механизмы и регуляцию этого процесса. Проницаемость стенки канальцев для ионов и воды определяется свойствами не только мембран клеток, но и зоны плотного соединения, где клетки контактируют друг с другом. Оба этих элемента существенно различаются в разных отделах нефрона. Через апикальную мембрану клетки натрий входит в цитоплазму пассивно по градиенту электрохимического потенциала, так как внутренняя поверхность клетки электроотрицательна по отношению к канальцевой жидкости. Далее натрий движется по цитоплазме к базальной и боковым частям клетки, где находятся натриевые насосы. В этих клетках натриевым насосом служит активируемая ионами Na+ и K+, зависимая от Mg+ аденозинтрифосфатаза (Na+, K+-АТФаза). Этот фермент, используя энергию АТФ, обеспечивает перенос из клетки ионов натрия и поступление в нее ионов калия. Ингибиторами этого фермента являются сердечные гликозиды (например, уабаин, строфантин К и др.), полностью прекращающие активную реабсорбцию натрия клетками проксимального канальца.
Важнейшее значение в осуществлении функциональной способности проксимального канальца имеет высокопроницаемая для некоторых ионов и воды зона клеточных контактов. Через нее происходят пассивная реабсорбция хлоридов и движение воды по осмотическому градиенту. Полагают, что регуляция скорости всасывания жидкости по межклеточным промежуткам происходит под влиянием таких физических сил, как соотношение между уровнем гидростатического давления в почечных артериях, венах и мочеточнике, величина онкотического давления в околоканальцевых капиллярах. Проницаемость межклеточных промежутков не является строго постоянной величиной, она может меняться при ряде физиологических состояний.
В тонком нисходящем отделе петли Генле не происходит сколько-нибудь существенной реабсорбции натрия и хлора. Особенностью этого канальца по сравнению с тонким восходящим отделом петли Генле является высокая проницаемость для воды. Тонкий нисходящий отдел петли характеризуется низкой проницаемостью для натрия, а восходящий, напротив, — высокой.
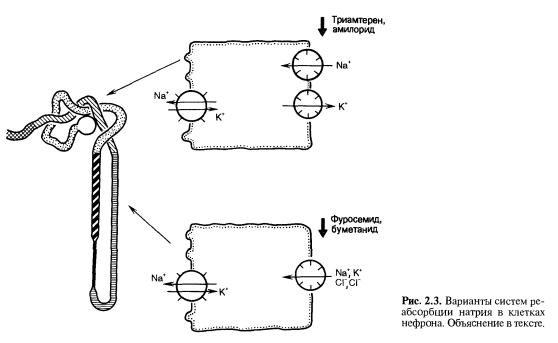
Пройдя по тонкому отделу петли Генле, жидкость поступает в толстый восходящий отдел петли. Стенка этого канальца всегда имеет низкую проницаемость для воды. Особенность клеток этого канальца состоит в том, что в них функционирует иной молекулярный механизм реабсорбции хлора и натрия из просвета канальца. В люминальной мембране расположена система переноса в клетку ионов Na+, K+ и двух ионов Cl-; эта макромолекула контранспортера инактивируется при наличии с внешней поверхности люминальной мембраны (т.е. в просвете канальца) таких веществ, как
фуросемид, буметанид, этакриновая кислота (рис. 2.3). С клинической точки зрения важно, что эти диуретики действуют из просвета канальца, они неэффективны при добавлении к околоканальцевой жидкости. Диуретики поступают в просвет нефрона при фильтрации и секреции в проксимальном канальце, с током мочи достигают толстого восходящего отдела петли Генле, прекращают реабсорбцию хлора и препятствуют всасыванию натрия в этом участке канальца.
Толстый восходящий отдел петли Генле переходит в часть дистального канальца, достигающую области macula densa, за которой следует дистальный извитой каналец. Этот отдел нефрона также обладает очень низкой проницаемостью для воды. Ведущим механизмом реабсорбции солей в этом канальце является
натриевый насос, который обеспечивает реабсорбцию натрия против высокого электрохимического градиента. Особенность реабсорбции натрия в этом отделе состоит в том, что хотя здесь может всосаться лишь 10 % профильтровавшегося натрия и скорость реабсорбции меньше, чем в проксимальном канальце, однако создается большой концентрационный градиент; концентрация натрия и хлора в просвете может снижаться до 30—40 ммоль/л. Реабсорбция хлора происходит вслед за реабсорбцией натрия и пассивно.
Связующий отдел соединяет дистальный сегмент нефрона с начальными отделами собирательных трубок, которые являются одними из важнейших структур почки, тонко и точно реагирующих на действие гормонов и приспосабливающих работу почки к потребности организма. В этих частях канальца основой реабсорбции натрия служит иной молекулярный механизм, не представленный в предшествующих отделах канальца. В люминальной мембране клеток имеются натриевые каналы, ингибируемые амилоридом и триамтереном.
Эти блокаторы натриевых каналов, являющиеся диуретиками, действуют из просвета нефрона. Они связываются с натриевыми каналами апикальной мембраны, которые обеспечивают вход натрия в клетку; когда канал блокирован амилоридом, натрий не может реабсорбироваться и экскретируется с мочой. В обычных условиях натрий, вошедший в клетку по натриевому каналу, движется по цитоплазме к базальным плазматическим мембранам и переносится во внеклеточную жидкость Na+-, K+-АТФазой. Стенка связующих канальцев и собирательных трубок может быть не только водонепроницаемой, но и высокопроницаемой для воды в присутствии АДГ. Именно в этом отделе канальцев действует АДГ. Транспорт натрия в этих клетках и дистальном извитом канальце увеличивается под влиянием альдостерона. В этих канальцах действует верошпирон, являющийся конкурентным ингибитором альдостерона. Верошпирон снижает реабсорбцию натрия, препятствуя связыванию в клетке альдостерона.
Кортикальные отделы собирательных трубок переходят в собирательные трубки, проходящие по мозговому веществу почки. Основная особенность их функции состоит в том, что они способны активно реабсорбировать совсем небольшие количества натрия, но могут создавать очень высокий концентрационный градиент. Стенка этих канальцев обладает низкой проницаемостью для солей и регулируемой проницаемостью для воды.
Раньше полагали, что реабсорбция жидкости в проксимальном канальце является облигатной, постоянной по величине и нерегулируемой в отличие от факультативной реабсорбции в дистальном сегменте канальцев, которая меняется при разных функциональных состояниях организма. Оказалось, что под влиянием импульсов, передающихся к почечным канальцам по эфферентным нервам, при действии ряда физиологически активных веществ меняется всасывание ионов и жидкости в проксимальном канальце. Это выявляется при увеличении объема крови и внеклеточной жидкости, когда уменьшение реабсорбции ионов и воды в проксимальном канальце способствует усилению их экскреции и нормализации водно-солевого обмена.

Важное значение в функции нефрона имеет возможность адаптивного изменения всасывания веществ в последующих отделах нефрона при нарушении их реабсорбции в вышележащих частях канальцев. Так, в некоторых случаях при уменьшении реабсорбции натрия в проксимальном сегменте нефрона он всасывается в восходящем отделе петли Генле (рис. 2.4). Эти данные имеют большое значение при оценке причин слабой эффективности фармакотерапии, направленной на усиление выведения определенных ионов почкой, при анализе механизма функции нефрона у больных с синдромом Фанкони и действии некоторых токсичных веществ на почку, когда отмечается усиление экскреции одних компонентов ультрафильтрата при нормальных остальных значениях состава мочи.
Для понимания особенностей выделения ионов натрия и хлора почкой, возможной резистентности отеков к действию диуретиков существенное значение имеют представленные выше данные об одновременной работе в клетках разных отделов почечных канальцев нескольких типов мембранных систем переноса натрия — натриевых каналов, чувствительных к амилориду, сопряженного переноса натрия, калия и хлора, блокируемого фуросемидом, а также ряда ионообменных систем, в которых натрий меняется на ионы калия, кальция, протоны. Оказалось, что и транспорт органических веществ (глюкоза, аминокислоты) сопряжен с переносом натрия. Такое большое разнообразие систем транспорта натрия в нефроне требует учета их роли при различных патологических состояниях почки. He меньшее значение приобретает выявление механизмов регуляции этих систем.
Регуляция выделения натрия почкой зависит от влияния на почку нескольких одновременно действующих на нее стимулов, поступающих по эфферентным нервам. Наиболее глубоко изучен механизм клеточного действия альдостерона, описание которого позволит глубже понять основные способы регуляции транспорта неорганических веществ в почке.
Альдостерон, введенный в кровь, увеличивает реабсорбцию натрия после продолжительного латентного периода, равного 45—120 мин. Этот гормон через перитубулярную плазматическую мембрану проникает в клетку и в цитоплазме связывается со стереоспецифичным для него белком. Этот процесс занимает 30—45 мин, после чего комплекс альдостерон — рецептор переносится в ядро и взаимодействует с акцепторными для него участками ядерно-го хроматина. Это вызывает транскрипцию гена: активация участка ДНК способствует синтезу РНК — посредника, который переходит из ядра в цитоплазму и стимулирует в рибосомах синтез нового белка. Существует ряд гипотез, объясняющих точку приложения в клетке этого белка и механизм, с помощью которого он увеличивает транспорт натрия. Согласно одной из гипотез, этот белок является компонентом натриевого канала, облегчая вход натрия в клетку через апикальную мембрану. Другая гипотеза основное значение придает синтезу макромолекул Na+-, К+-АТФазы. Высказывается также мысль о возможности действия этого белка не на один, а на два компонента системы — вход и активное выведение натрия из клетки, что автоматически усиливает энерготраты и приводит к возрастанию образования АТФ. Понимание механизма действия альдостерона имеет существенное значение для клиники: становятся понятными способ действия, причина длительного латентного периода и сущность влияния диуретиков, являющихся конкурентными антагонистами альдостерона (верошпирон). Эти вещества угнетают связывание альдостерона с рецепторными белками, находящимися в цитоплазме.
По новым данным, основным: местом действия альдостерона в нефроне являются самые конечные отделы дистального извитого канальца и начальные отделы собирательных трубок. В этих же клетках действуют и спиронолактоны. Описанные выше данные о механизме клеточного влияния альдостерона касаются только его влияния на реабсорбцию натрия; вызываемое гормоном: усиление секреции калия не опосредовано генетическим аппаратом клетки.
Калий. Основное количество калия в организме находится в клетках; во внеклеточной жидкости, включая плазму крови, содержится лишь 2 % калия — около 65 ммоль. С пищей человек потребляет в день 25—150 ммоль калия, т.е. во всей внеклеточной жидкости содержится в среднем почти столько калия, сколько поступает с пищей и выводится почкой. Таким образом, становится очевидной роль почки в гомеостазе калия, концентрация которого должна поддерживаться в особо узких пределах, поскольку гипер- и гипокалиемия могут вызвать тяжелые нарушения в работе клеток различных органов и систем.
Калий свободно фильтруется в клубочках, в сутки в просвет канальцев поступает 700—800 ммоль калия, который реабсорбируется клетками проксимального и дистального сегментов нефрона. Экскреция калия составляет обычно около 10 % профильтровавшегося количества, но механизм выделения калия почкой иной, чем натрия. Профильтровавшийся калий почти полностью реабсорбируется в проксимальном сегменте и петле Генле, а выделение с мочой зависит от его секреции клетками дистального извитого канальца и собирательных трубок. При избыточном поступлении калия в организм его реабсорбция в проксимальных канальцах не снижается, но резко усиливается секреция в конечных отделах почечных канальцев. Особое значение секреторный механизм приобретает при уменьшении объема клубочковой фильтрации; количество секретируемого калия в ряде случаев может превышать количество всего калия, поступающего в нефрон при фильтрации в клубочках.
В одной и той же клетке дистального извитого канальца и собирательных трубок, по-видимому, сосуществуют системы реабсорбции и секреции калия. При дефиците калия они обеспечивают максимальное извлечение калия из канальцевой жидкости, а при избытке — его секрецию.
Калий, секретируемый клеткой, транспортируется вначале в цитоплазму из внеклеточной жидкости Na/K-насосом, находящимся в базальной плазматической мембране. Этот насос обеспечивает удаление из клетки натрия и поступление в нее калия, что создает высокую внутриклеточную концентрацию калия. Для секреции необходимо, чтобы возросла проницаемость .номинальной плазматической мембраны для калия и градиент потенциала на этой мембране благоприятствовал его секреции. Увеличение отрицательного электрического заряда просвета дистального канальца при введении сульфата натрия способствует секреции калия. Альдосте-рон является одним из факторов, увеличивающих проницаемость апикальной мембраны для калия и повышающих его секрецию. Таким образом, секреция калия через мембрану клетки со стороны просвета канальца является пассивным процессом, происходящим по концентрационному градиенту и зависящим от проницаемости мембраны для калия и величины электрического потенциала на этой мембране.
Усиление секреции калия под влиянием альдостерона связано не только с действием последнего на проницаемость мембраны для калия, но и с увеличением поступления калия в клетку вследствие усиления работы Na/K-насоса. Другим важным фактором регуляции транспорта калия в канальцах является инсулин, уменьшающий экскрецию калия и повышающий толерантность организма к введению его солей. Большое влияние на уровень выделения калия оказывает КОС. Алкалоз сопровождается увеличением выделения калия почкой, а ацидоз приводит к уменьшению калийуреза.
Кальций. Общее количество кальция в организме велико — около 2 % массы тела, поскольку этот элемент является одним из основных компонентов скелета.
Почки и кость играют главную роль в поддержании стабильного уровня кальция в крови, что исключительно важно в связи с ролью кальция в большом числе физиологических процессов: он участвует в секреции, оплодотворении, мембранной проницаемости, свертывании крови, мышечном сокращении; ионы кальция оказались внутриклеточными мессенджерами. В сутки потребление кальция составляет около 1 г; 0,8 г выделяется кишечником, почки экскретируют от 0,1 до 0,3 г (2,5—7,5 ммоль/сут).
В клубочках фильтруется кальций, ионизированный и находящийся в виде низкомолекулярных комплексов, что составляет до 11 г/сут. Более 50 % профильтровавшегося кальция реабсорбируется в проксимальном канальце, значительная реабсорбция кальция происходит в толстом восходящем отделе петли Генле (20—25 %), дистальном извитом канальце (5—10 %) и собирательных трубках (0,5—1 %). С мочой экскретируется кальций, «избежавший» реабсорбции; секреции кальция в почке у человека не происходит.
Механизм реабсорбции кальция представляется следующим образом. Общее внутриклеточное содержание кальция составляет около 0,3 ммоль, т.е. меньше, чем в плазме крови. Однако в самой цитоплазме находится лишь около 0,01 % свободного кальция (около 10в-7 М) от общего количества, в основном он сосредоточен в эндоплазматической сети и митохондриях. Если во внеклеточной жидкости и ультрафильтрате, поступающем в каналец, концентрация кальция составляет около 1,3 ммоль/л, то в цитоплазме клетки она соответственно в 1000— 10 000 раз меньше. Выделение кальция из клетки при реабсорбции происходит активно через базальную плазматическую мембрану с помощью кальциевого насоса (Са2+-АТФаза). Другим механизмом в этой же мембране служит Nа+/Са2+-обменник, обеспечивающий удаление ионов Ca2+ из клетки в обмен на поступающие в нее ионы натрия. Условием работы этого молекулярного устройства служит непрерывное выведение из клетки ионов натрия при участии находящейся рядом в мембране Na-, К-АТФазы. В клетке почечного канальца нужно иметь особенно эффективную систему стабилизации содержания Ca2+, так как он непрерывно поступает через люминальную мембрану, а ослабление транспорта в кровь не только нарушило бы баланс кальция в организме, но и повлекло бы патологические изменения в самой клетке нефрона. В базальной плазматической мембране имеется также Ca2+/Na2+-обменник; временное перераспределение кальция внутри клетки обеспечивают эндоплазматическая сеть и отчасти митохондрии.
Среди гормонов, регулирующих транспорт кальция в почке, наибольшее значение имеет паратгормон. После удаления паращитовидных желез резко возрастает кальцийурез, несмотря на уменьшение количества фильтруемого кальция из-за гипокальциемии. Введение паратгормона уменьшает выделение кальция почкой. В проксимальном канальце паратгормон уменьшает реабсорбцию кальция, однако при этом снижается его экскреция почкой вследствие стимулируемого этим гормоном всасывания кальция в дистальном сегменте нефрона и собирательных трубках.
В противоположность паратгормону тирокальцитонин вызывает увеличение экскреции кальция почкой. Активная форма витамина D3 повышает реабсорбцию кальция, по-видимому, в проксимальном канальце нефрона. Гормон роста способствует усилению кальцийуреза, хотя неясно, обусловлено ли это прямым действием гормона на клетки нефрона или опосредованным, связанным с хроническим увеличением объема внеклеточной жидкости. Повышенное выделение кальция с мочой в условиях действия на почку паратгормона позволяет видеть в этом одну из причин частого появления мочекаменной болезни у больных акромегалией.
Рецепторы паратгормона в почке найдены в базальной и латеральной мембранах клеток проксимального сегмента нефрона, корковых частях толстого восходящего отдела петли Генле и начальных отделах собирательных трубок. После стимуляции рецепторов этим гормоном активируется фермент аденилатциклаза, благодаря чему увеличивается внутриклеточная концентрация цАТФ и меняется транспорт кальция и фосфатов.
Магний. В норме у человека в плазме крови содержится около 0,8—1 ммоль*л-1 магния, в просвет нефрона при фильтрации в минуту поступает около 0,064 ммоль ионизированного магния и 0,01 ммоль в виде комплексов; остальной магний остается в плазме, так как он связан с белками. Выделение с мочой колеблется от 3 до 5 ммоль/сут. Исследования методом микропункции выявили значительные различия в уровне реабсорбции магния в проксимальном канальце — от 20 до 60 %; большие количества магния всасываются клетками толстого восходящего отдела петли Генле. Реабсорбция магния является активным процессом и ограничена величиной максимального канальцевого транспорта (TmMg). Гипермагниемия приводит к усилению экскреции магния почкой и может сопровождаться преходящей гиперкальциурией. При нормальном уровне клубочковой фильтрации почка быстро и эффективно справляется с повышением уровня магния в крови, предотвращая гипермагниемию, поэтому клиницисту чаще приходится встречаться с проявлениями гипомагниемии. Повышенный уровень магния в крови может наблюдаться при резком снижении скорости клубочковой фильтрации.
До сих пор не установлены специфические факторы, регулирующие выделение магния почкой, хотя гомеостаз магния поддерживается исключительно точно. Обнаружена система рефлекторной регуляции выделения магния почкой. Скорость экскреции магния возрастает при остром увеличении объема внеклеточной жидкости, при инъекции больших доз тирокальцитонина и вазопрессина; паратгормон уменьшает выделение магния. Однако гиперпа-ратиреоидизм у ряда больных сопровождается гипомагниемией и отрицательным балансом магния, что, возможно, обусловлено более выраженным действием гиперкальциемии на почку, чем прямым влиянием паратгормона. Гиперкальциемия вызывает увеличение экскреции кальция и магния почкой.
Фосфор. Почки играют ключевую роль в поддержании постоянства концентрации фосфатов в жидкостях внутренней среды. В плазме крови фосфаты представлены в виде свободных ионов (около 80 %) и связанных с белками. При pH 7,4 в плазме крови почти 80 % фосфатов существует в двухвалентной форме HPO4-, 20 % составляют H2PO4-. При нормальном уровне клубочковой фильтрации 125 мл/мин и концентра-
ции фосфатов 1,14 ммоль*л-1 в клубочках фильтруется около 0,132 ммоль*мин-1 и экспедируется в среднем 10 % этого количества, 90 % реабсорбируется в канальцах. Выделение почкой обычно колеблется в пределах 13 — 42 ммоль/сут. Наиболее интенсивно всасываются фосфаты в начальных частях проксимального канальца, где реабсорбируется 60—70 % фильтруемых фосфатов, в петле Генле до дистального извитого канальца реабсорбируется лишь 5— 10 % фосфатов, в дистальном канальце и собирательных трубках — 10—25 %. Такова картина обработки фосфатов в нормальных условиях; если резко снижена проксимальная реабсорбция фосфатов, то полнее начинает использоваться большая мощность транспортной системы дистального сегмента нефрона, который может в этих условиях предотвращать фосфатурию.
Основным фактором регуляции реабсорбции фосфатов служит паратгормон. При гиперпара-тиреоидизме резко увеличена экскреция неорганических фосфатов, дефицит паратгормона проявляется в стимуляции реабсорбции фосфатов. Транспорт фосфатов ограничен TmPO4, введение паратгормона резко уменьшает величину TmPO4 в проксимальном и дистальном сегментах нефрона. Роль этого гормона заключается в регуляции транспорта фосфатов в различных отделах нефрона. Механизм фосфатурического действия паратгормона основан на увеличении внутри клеток, чувствительных к действию этого гормона, цАМФ, который уменьшает реабсорбцию фосфатов.
В проксимальном канальце 1,25 (OH)2D3 стимулирует реабсорбцию фосфатов. Важно отметить, что одной из функций клеток коры почки является превращение неактивной формы циркулирующего в крови витамина D3 в активные метаболиты. Другим биологически активным веществом, усиливающим реабсорбцию фосфатов, является гормон роста. В почечной ткани найдены химические рецепторы тирокальцитонина, однако его реальное значение в регуляции транспорта фосфатов пока не установлено. Фосфатурическое действие характерно для диуретиков, эффект которых локализован в проксимальном канальце.
Сульфат. Концентрация сульфата в плазме крови составляет 0,6+0,2 ммоль/л. Профильтровавшиеся в клубочках сульфаты реабсорбируются в канальцах системой с TmSO4, максимальная реабсорбция которой равна 0,04—0,12 ммоль*мин1. При введении сульфатов в кровь быстро насыщается система реабсорбции, и избыток сульфатов экскретируется почкой. Максимальная способность к реабсорбции сульфатов снижается после введения в кровь фосфатов, ацетата, тиосульфата. Величина максимальной реабсорбции сульфатов возрастает после введения гормона роста и глюкокортикоидов, но не дезоксикортикостерона. Почти в 2 раза увеличивался TmSO4 у больных акромегалией в активном периоде болезни. Возможно, что действие гормона роста носит не специфический характер, а обусловлено возрастанием клубочковой фильтрации и реабсорбции всех основных компонентов ультрафильтрата.







 23.01.2018
23.01.2018