 Главная
Главная
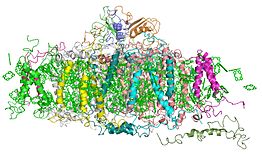
Фотосистема I (первая фотосистема, фотосистема один, ФСI), или пластоцианин-ферредоксин-оксидоредуктаза — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.
История открытия
Нециклический транспорт электронов начинается с того, что марганцевый кластер фотосистемы II окисляет воду, восстанавливая пул пластохинонов. Далее цитохром b6f-комплекс окисляет пластохиноны, а электрон через пластоцианин передаётся на фотосистему I, где используется для синтеза НАДФН. Нарушение формальной логики в названиях фотосистем связано с тем, что фотосистема I была открыта раньше, чем фотосистема II.
Первые данные указывающие на существование ФСI появились в 1950-х годах, но в то время никто ещё не мог оценить значимость этих открытий. Представление о существовании в хлоропластах двух фотосистем возникло в уже в 1940-е годы на основание опытов лаборатории Р. Эмерсона, обнаружившего эффект падения квантового выхода фотосинтеза при освещении хлоропластов монохроматическим красным светом (λ>680 нм), возбуждающим только ФСI, и эффект усиления квантового выхода при добавлении подсветки с длиной волны около 650 нм, которая возбуждала ФСII (так называемый эффект Эмерсона). Также следует упомянуть открытый Комоннером в 1956 году светоиндуцируемый ЭПР сигнал, который был назван сигналом I. По чистой случайности сигнал I и сигнал II, исходили от ФСI и ФСII соответственно. Лишь в 1960 году Луис Дюйзенс предложил концепцию фотосистемы I и фотосистемы II, и в том же году Фэй Бендалл и Роберт Хилл организовали результаты предыдущих открытий в стройную теорию последовательных реакций фотосинтеза. Гипотеза Хилла и Бендалля была позднее подтверждена в экспериментах Дюйзенса и Витта в 1961 году.
После этого начались систематические попытки физического выделения фотосистемы I, определения её трёхмерной структуры и тонкого строения. В 1966 году начался бум исследований в этой области: Андерсон и Бордман подвергали мембраны хлоропластов воздействию ультразвука с последующей обработкой дигитонином, Вернон использовал тритон X-100, а Огава — додецилсульфат. Однако, первые полученные экстракты содержали примеси светосособирающих комплексов, а также цитохромов f и b6. Потребовалось много времени, чтобы разобраться, что полученные экстракты были смесью.
В 1968 году Рид и Клайтон смогли выделить реакционный центр фотосистемы I из пурпурных бактерий, что значительно подстегнуло исследования оксигенного фотосинтеза. Однако, открытым оставался вопрос: что из выделенного являлось истинным реакционным центром, что антенными комплексами, а что дополнительными субъединицами. Долгое время эффективное выделение реакционного центра фотосистемы I оставалось неразрешённой проблемой. В конце-концов оказалось, что легче всего это сделать у цианобактерий, поскольку у них отсутствовали интегрированные в мембрану внешние антенны. После многочисленных попыток с разными видами, выяснилось, что самыми многообещающими в этом отношение видами являются представителями Synechocystis и Synechococcus, поскольку фотосистема I, выделенная из Thermosynechococcus elongatus давала очень стабильный реакционный центр, пригодный для кристаллизации и исследования методом рентгеноструктурного анализ.
Отличия от фотосистемы II
Основная функция фотосистемы II — генерация сильного окислителя, который инициирует окисление воды и передачу её электронов на мембранный переносчик. Основная функция фотосистемы I — насытить эти низкоуровневые электроны энергией, чтобы с их помощью осуществить восстановление НАДФ+. Поскольку энергия суммарного процесса слишком велика, чтобы осуществить его в рамках одного реакционного центра, в ходе эволюции появились две фотосистемы, которые раздельно осуществляют разные части этой реакции. Их специфические функции и определяют особенности их строения. Так, фотосистема I — симметрична, то есть в ней работают две ветви электронного транспорта, что делает его значительно более быстрым, в то время как фотосистема II — асимметрична и обладает только одной рабочей ветвью, что замедляет транспорт электронов, но делает его более управляемым. Обе фотосистемы значительно отличаются по строению антенн, дополнительных субъединиц, способов регуляции и своему положению в мембране. Так, фотосистема I обладает интегральной антенной, хлорофиллы которой расположены непосредственно на главных белках комплекса — А и B, в то время как у фотосистемы II они вынесены на внешние белки CP47 и CP43. По количеству дополнительных малых регуляторных субъединиц ФС II значительно превосходит ФС I, что связано с необходимостью тонкой регуляции процесса окисления воды, который потенциально крайне опасен для клетки. Этим же объясняется неоднородное распределение фотосистем в мембране тилакоида: в то время как ФС I располагается преимущественно в области маргинальных, торцевых и стромальных мембран, ФС II практически полностью находится в области спаренных мембран, что обеспечивает клетке дополнительную защиту от продуцируемых ею активных форм кислорода.
Главное отличие фотосистемы II от фотосистемы I — это наличие большого обращённого в люмен домена, который содержит марганцевый кластер и окружающие его защитные белки. Именно здесь происходит процесс фотохимического окисления воды, сопровождаемый выделением кислорода и протонов.
Структурная организация фотосистемы I
Фотосистема I состоит из следующих белковых субъединиц и кофакторов:
Основная функция ФСI — передача энергии света на электрон, перенос электрона от пластоцианина к ферредоксину. ФСI содержит свыше 110 кофакторов, значительно больше чем фотосистема II. Каждый из этих компонентов имеет широкий спектр функций. Основные компоненты электрон-транспортной цепи ФСI — главный донор возбуждённых электронов П700 (хлорофильный димер) и пять переносчиков: A0 (хлорофилл а), A1 (филлохинон) и три Fe4S4 железосерных кластера: Fx, Fa, и Fb.
Структурно ФСI представляет собой гетеродимер двух интегральных белковых комплексов — А и В (у всех растений кодируются хлоропластными генами PsaA и PsaB). Белки А и В присоединяют димер Р700, по одной молекуле мономера хлорофилла а (Хл695) — первичного акцептора электронов А0, по одному дополнительному хлорофиллу а и по одной молекуле филлохинона (А1). Два набора дополнительных хлорофиллов а, первичных акцепторов электрона и филлохинонов формируют две почти симметричные ветви транспорта электронов от Р700 к Fx. В отличие от реакционных центров зелёных и пурпурных бактерий и ФСII, где из двух ветвей функционирует лишь одна, в ФСI активны обе ветви электронного транспорта, хотя они и не идентичны.Белок А гомологичен белкам D1+СP43 (молекулярная масса белка А соответствует сумме молекулярных масс белков D1 и СP43) из фотосистемы II, а белок В гомологичен белкам D2+CP47 соответственно.
Обе субъединицы содержат 11 трансмембранных сегментов. Железосодержащий кластер Fx связан четырьмя цистеинами, два из которых находятся на субъединице А, и ещё два на субъединице В. В обоих белках цистеины находятся на проксимальном конце, в петле между девятым и десятым трансмембранными сегментами. По всей вероятности, ниже цистеинов находится так называемый мотив лейциновой молнии, который вносит существенный вклад в димеризацию белков А и В. Конечные акцепторы электронов FA и FB находятся на субъединице С.
Следует особо подчеркнуть, что перенос электрона осуществляется в соответствии с термодинамическим потенциалом. Увеличение окислительно-восстановительных потенциалов в цепи акцепторов обеспечивает быстрое снижение энергии, что предотвращает возврат электрона к пигменту и бесполезную трату энергии электронного возбуждения. Благодаря этому энергия возбуждения эффективно используется для разделения зарядов.
Пластоцианин
Пластоцианин — маленький, подвижный белок с молекулярной массой около 10,5 кДа. К его центральному атому Сu присоединяются остатки цистеина и метионина, а сбоку его стабилизируют два остатка гистидина. При обратимой смене валентности Cu2+ ↔ Cu+1 пластоцианин либо поглощает один электрон, либо отдаёт его. Пластоцианин является аналогом цитохрома c, который выполняет схожую функцию в дыхательной цепи митохондрий.
Он принимает электрон от цитохром b6f-комплекса, окисляя цитохром f и перенося его непосредственно на реакционный центр П700 фотосистемы I. На внешней стороне белка имеется группа аминокислот, несущих отрицательный заряд. Предположительно, они связываются с положительно заряженным люминальным доменом субъединицы F, однако механизм связывания недостаточно изучен и остаётся неясным.
У некоторых водорослей и цианобактерий при недостатке меди в среде пластоцианин не образуется, вместо него синтезируется и выполняет его функции цитохром c-553.
- Пластоцианин (PC) отдаёт один электрон окисленному П700+ и восстанавливает его до исходного состояния:
Специальная пара П700
П700 (в английской литературе P700) представляет собой димер хлорофилла а и хлорофилла а‘ у которого кетоэфирная группа в V кольце находится в цис-положении относительно плоскости молекулы, с максимумом поглощения 700 нм. Наличие цис-кетоэфирной группы делает возможным образование димера из двух хлорофиллов посредством формирования водородных связей. П700 получает энергию от антенных комплексов и использует её, чтобы поднять электроны на более высокий уровень. Далее электрон в ходе окислительно-восстановительной реакции переходит на цепь переносчиков. В окисленном состоянии окислительно-восстановительный редокс-потенциал П700 составляет +0,52 В, а в фотовозбуждённом состоянии он становится −1,2 В, то есть формируется мощный восстановитель, обеспечивающий восстановление НАДФ+.
- В соответствии со следующим уравнением П700 поглощает квант света и переходит в фотовозбуждённое состояние, в результате чего один из его электронов переходит с основного подуровня S0 на первый синглетный подуровень S1:
Хлорофилл А0
A0 — первый акцептор электронов в фотосистеме I. Именно здесь происходит первичное фотохимическое разделение зарядов между фотовозбуждённым П700* и A0. Его максимум поглощения составляет 695 нм (Хл а695), что объясняется его взаимодействием с окружающими аминокислотными остатками. Его редокс-потенциал в восстановленном состоянии −1,1 В.
- Фотовозбуждённый П700* отдаёт один электрон хлорофиллу A0, в результате чего происходит разделение зарядов, и образуется первичная радикальная пара:
Филлохинон A1
Следующий акцептор — это Филлохинон A1, также известный как витамин K1. Он, как и хлорофилл, обладает фитольным хвостом, и приблизительно соответствует пластохинону QA фотосистемы II. Поглощая электрон, он образует семихинон-радикал, который восстанавливает Fx, передаёт его на Fb и далее на Fa.
A 0 − + A 1 → A 0 + A 1 ⋅ {displaystyle {mathsf {A_{0}^{-}+A_{1} ightarrow A_{0}+A_{1}cdot }}}Железосерные кластеры
Железосерные кластеры ФСI имеют форму куба с четырьмя атомами железа и четырьмя атомами серы, составляющими его восемь вершин. Все три кластера связаны с белками ФСI через остатки цистеина. Fx (Eо‘ = −0,70 В) окисляет восстановленный А1. Дальнейший транспорт осуществляют железосерные кластеры Fa, и Fb, характеризующиеся низкими окислительно-восстановительными потенциалами (-0,59 и −0,55 В соответственно). Множество экспериментов выявило несоответствие между разными теориями, описывающими расположение и работу железосерных кластеров. Однако большинство результатов позволяет сделать некоторые общие выводы. Во-первых, Fx, Fa, и Fb образуют треугольник, и Fa располагается ближе к Fx чем Fb. Во-вторых, транспорт электронов начинается с Fx через Fa к Fb, или же через Fa к Fb. До сих пор ведутся споры о том, какой из двух кластеров осуществляет перенос электрона на ферредоксин.
Ферредоксин
Ферредоксин — это водорастворимый белок с молекулярной массой 11 кДа и содержащий Fe2S2 центр. Примечательно, что он является одноэлектронной окислительно-восстановительной системой, то есть переносит только один электрон, полученный им от железосерных кластеров. Он восстанавливается ФСI на стромальной стороне мембраны и в восстановленном состоянии является сильным восстановителем (Eo‘ = —0,6 В), благодаря чему может быть переносчиком электронов для различных реакций, протекающих в хлоропласте. Так, ферредоксин поставляет электроны для восстановления нитритов, (нитритредуктаза) и ассимиляции серы (сульфитредуктаза) в хлоропласте. Также он поставляет электроны для фиксации атмосферного азота (нитрогеназа) у бактерий. Он восстанавливает тиоредоксин — низкомолекулярный серосодержащий белок, участвующий в редокс-регуляции хлоропластов, активируя ключевые ферменты цикла Кальвина. При нециклическом транспорте электронов ферредоксин взаимодействует с ферредоксин-НАДФ(+) редуктазой, которая восстанавливает НАДФ+ до НАДФН (Eo‘ = −0,32 В) в строме хлоропласта.
Светособирающий комплекс
ФСI и ССКIСветособирающие комплексы состоят из молекул хлорофилла а и b и каротиноидов, соединённых с белками. Эти пигменты, возбуждаясь, переносят энергию фотонов в реакционный центр фотосистемы по Фёрстеровскому механизму. В отличие от реакционного центра ФСI, светособирающие комплексы могут поглощать практически во всей области видимого спектра. Антенные комплексы подразделяют на внутренние, или интегральные, антенны, непосредственно присоединённые к комплексу фотосистемы, и периферические подвижные светособирающие комплексы (ССКI). Так, белки А и В присоединяют пигменты внутренней антенны ФСI: около 95 молекул хлорофилла а и 22 молекулы β-каротина, 5 из которых находятся в цис-конформации. В координации по крайней мере десяти хлорофиллов внутренней антенны принимают участие малые субъединицы J, K, L, M и X. Пигменты внутренней антенны располагаются в виде цилиндра, окружающего все компоненты электрон-транспортной цепи фотосистемы I. Это отличает ФСI от ФСII, где пигменты внутренней антенны расположены на отдельных белках CP43 и CP77. Внешний светособирающий комплекс ССКI (LHCI) содержит 80—120 молекул хлорофиллов а и b, каротиноиды и состоит из четырёх субъединиц: Lhca1, Lhca2, Lhca3 и Lhca4 — с молекулярными массами 17—24 кДа. Относительно недавно были открыты две дополнительные субъединицы, Lhca5 и Lhca6, однако их концентрация в мембране тилакоида крайне мала, а кодирующие их гены практически не экспрессируются.
Циклический транспорт электронов
При слишком сильном освещении и/или закрытых устьицах (голодание по СО2) происходит перевосстановление пула пластохинонов и, как следствие, перевосстановление пула НАДФ+. При недостатке СО2 НАДФН не может расходоваться в цикле Кальвина, а значит, не хватает субстрата для ферредоксин-НАДФ+-редуктазы. В конечном итоге это приводит к тому, что ФСI становится некуда сбрасывать возбуждённые электроны, а это в свою очередь может привести к повреждению фотосинтетического аппарата, окислению мембран и образованию активных форм кислорода. В этих условиях, чтобы предотвратить окислительный стресс и защититься от фотоповреждения, растения переходят к циклическому транспорту электронов. Полагают, что катализатором циклического транспорта является восстановленный ферредоксин.
Циклическое фотофосфорилирование
Вначале электрон неким образом перемещается от восстановленного ферредоксина на пул пластохинонов. Точный механизм этого процесса не известен. Полагают, что эту реакцию осуществляет особый фермент — ферредоксин-пластохинон-оксидоредуктаза. Затем от пластохинона через цитохром b6f-комплекс и пластоцианин электрон вновь попадает на ФСI. При этом происходит закачка протона в полость тилакоида и идёт синтез АТФ. В качестве наиболее вероятного кандидата на роль ферредоксин-пластохинон-оксидоредуктазы в последнее время рассматривают ферредоксин-НАДФ+-редуктазу, которая может образовывать комплекс с цитохром-b6f-комплексом. Предположительно она может переносить электроны с ферредоксины напрямую на убихинон, связанный цитохром-b6f-комплексом через специальный гем cn. Большое количество данных также говорит в пользу образования суперкомплекса из цитохром-b6f-комплекса, ФСI, ферредоксин-НАДФ+-редуктазы и трансмембранного белка PGRL1. Образование и распад такого комплекса, как полагают, переключает режим потока электрона с нециклического на циклический и обратно.
Ещё один фермент, возможно принимающей участие а этом процесс — это НАДН-дегидрогеназный комплекс хлоропластов, аналогичный НАДH-дегидрогеназному комплексу митохондрий и гомологичный бактериальному комплексу I. Он окисляет ферредоксин и сбрасывает электроны на пластохинон, предотвращая окислительный стресс. НАДН-дегидрогеназный комплекс хлоропластов образует суперкомплекс с двумя ФСI при помощи белков Lhca5 и Lhca6. Протонный градиент, созданный в результате циклического фотофосфорилирования на мембране тилакоида, используется белками-переносчиками для встраивания в мембрану приходящих из стромы белков.
Псевдоциклический транспорт
При очень активном восстановлении пула ферредоксинов происходит сброс их электронов на О2 с образованием H2O (так называемая реакция Мелера). Он сходен с циклическим транспортом тем, что не синтезируется НАДФН, а только АТФ. Однако в условиях реакции Мелера соотношение АТФ/АДФ очень велико, так что имеющегося количества АДФ не достаточно для синтеза АТФ, и, как следствие, на мембране тилакоида создаётся очень высокий протонный градиент. В результате реакции происходит образование супероксид-анион-радикала O2-·, который превращается в O2 и H2O2 под воздействием фермента супероксиддисмутазы, а перекись превращается в воду ферментом аскорбатпероксидазой.
F d r e d + O 2 → F d o x + O 2 − ⋅ {displaystyle {mathsf {Fd_{red}+O_{2} ightarrow Fd_{ox}+O_{2}^{-}cdot }}}Ещё один фермент, участвующий в псевдоциклическом транспорте, это терминальная оксидаза хлоропластов, гомологичная альтернативной оксидазе растительных митохондрий. Она окисляет пул пластохинонов с участием кислорода, образуя воду и рассеивая энергию в форме тепла.
Локализация в мембране тилакоида
Фотосистема I находится в стромальных тилакоидах (32 %), а также в маргинальных (36 %) и торцевых (32 %) областях гран. Такое расположение обусловлено плотностью её поверхностного заряда и силами электростатического отталкивания с другими комплексами.
У цианобактерий и прохлорофитов фотосистема I способна образовывать тримеры. Это способствует увеличению спектра поглощения на больших глубинах, а также более эффективному перераспределению энергии возбуждения и защите от фотоповреждений. Следует отметить, что у эукариот фотосистема I утратила эту способность благодаря наличию субъединицы H, а также мутации в субъединице L. Вместо тримерезации у эукариот она при помощи субъединиц L и G взаимодействует с большими мембранными светособирающими комплексами, которых нет у прокариот.
Белок Ycf4
Трансмембранный белок Ycf4, обнаруженный в мембране тилакоида, жизненно необходим для функционирования фотосистемы I. Он участвует в сборке компонентов комплекса, без него фотосинтез становится неэффективным.
Зелёные серобактерии и эволюция ФСI
Молекулярно-биологические данные говорят в пользу того, что ФСI, вероятно, эволюционировала из фотосистемы зелёных серобактерий. Реакционные центры зелёных серобактерий, цианобактерий, водорослей и высших растений различаются, однако домены, выполняющие аналогичные функции, имеют схожее строение. Так, во всех трёх системах окислительно-восстановительный потенциал достаточен для восстановления ферредоксина. Все три электронотранспортные цепи содержат железо-серные белки. И наконец, все три фотосистемы представляют собой димер двух гидрофобных белков, на которых закреплены редокс-центры и пигменты интегральной антенны. В свою очередь, фотосистема зелёных серобактерий содержит те же самые кофакторы, что и электронотранспортная цепь фотосистемы I.
Галерея
-
Положение хлорофиллов и кофакторов в фотосистеме I.
-
Тример фотосистемы I
-
ЭТЦ фотосистемы I
-
Фотосистемы I и реакционный центр бактерии.
-
Модель фотосистемы I.